Après avoir parcouru certaines publications et/ou sites sur Internet et, plus encore, ayant découvert les interventions de quelques uns sur les réseaux sociaux, le moins que je puisse en conclure est que la notion même de ce que représente la sélection naturelle dans le processus général de l’Évolution est plutôt mal connue. Lorsqu’elle n’est pas totalement dénaturée… Il parait donc utile de revenir aujourd’hui (et bien que nous l’ayons déjà longuement évoquée sur ce blog) sur ce qu’est vraiment la sélection naturelle dans la grande machinerie de l’Évolution.
la théorie de l’Évolution est un ensemble de
lois validées par l’expérience
Précisons tout d’abord que la théorie de l’Évolution est bel et bien comme le dit le dictionnaire une théorie, c’est-à-dire « un ensemble organisé de principes, de règles, de lois scientifiques visant à décrire et à expliquer un ensemble de faits » et non pas comme le souhaiteraient les créationnistes de tous poils une sorte de juxtaposition d’hypothèses plus ou moins justifiées selon la bonne volonté de leurs auteurs. La théorie de l’Évolution revue et corrigée par ses aspects les plus modernes est à ce jour la meilleure explication possible à la manière dont est organisée la vie sur Terre. Comme la théorie de la relativité générale est pour le moment la plus susceptible d’expliquer l’univers qui nous entoure. Ces précisions étant faites, essayons d’aller un peu plus avant.
Dès la publication de son ouvrage, Charles Darwin (illustration) a bien expliqué que ce qui lui paraissait être le but de l’évolution des espèces était leur maintien le plus longtemps possible dans la sphère du vivant et donc leur adaptation continue au fur et à mesure de la survenue des inévitables changements du milieu au sein duquel elles vivent.
maintien le plus longtemps possible dans la sphère du vivant et donc leur adaptation continue au fur et à mesure de la survenue des inévitables changements du milieu au sein duquel elles vivent.
Il est bon de rappeler que le scientifique anglais fit part de sa découverte à un moment où la science était encore balbutiante dans de nombreux domaines. La génétique, par exemple, était totalement inconnue (ce qui obligea d’ailleurs Darwin à recourir partiellement à une explication par la transmission des caractères acquis !). De même, l’embryologie, la biologie, la génétique des populations, etc. n’existaient pas non plus ou si peu : Charles Darwin n’en eut que plus de mérite à bâtir une théorie que ces disciplines alors ignorées allaient largement valider par la suite.
Un des mécanismes fondamentaux de l’Évolution des êtres vivants est la sélection naturelle, à savoir la survie du plus apte, de celui qui présente le degré de résistance le plus élevé aux changements de son environnement, celui qui, en somme, possède sur tous les autres un avantage évolutif qui lui permet de survivre là où ses concurrents ne le peuvent pas.
Où agit réellement la sélection naturelle ?
Son action peut se situer à différents niveaux dont certains, parfois, sont peu intuitifs.
* l’individu : pour Darwin, c’est à travers l’individu que se signale cette faculté supérieure d’adaptation ; pour pouvoir se reproduire, en effet, un sujet doit tout d’abord atteindre l’âge de la maturité afin de rencontrer le ou la partenaire qui lui permettra de se reproduire puis il devra avoir la descendance la plus nombreuse possible, susceptible d’échapper ainsi aux « aléas » de l’existence. S’il est porteur d’un avantage significatif, il ne lui en sera que plus facile de « distancer » les autres. Cette approche de la sélection dite naturelle est longtemps restée ultra-dominante avant que d’autres évolutionnistes ne cherchent à l’étendre à d’autres niveaux.
* le groupe : pour certains évolutionnistes, l’individu, s’il est important, n’est pas seul dans la sélection : il faut également regarder le groupe auquel il appartient. Effectivement, dans certains cas plus nombreux qu’on l’imagine, il existe des sortes de collaborations innées comme nous avons pu le voir dans le sujet qui a été consacré à « l’intelligence animale collective ». En pareil cas, l’association de plusieurs individus leur permet d’être bien plus performants dans leur survie globale que s’ils avaient dû rester autonomes. Un exemple bien connu est celui des fourmis : seul, un  individu ne peut rien mais avec l’aide de ses congénères, il appartient alors à une force tout à fait redoutable, susceptible non seulement de s’attaquer à des ennemis en apparence bien plus importants mais également capable d’actions spectaculaires et parfois difficilement imaginables. D’autres insectes sociaux (abeilles, termites, araignées sociales, etc.) décuplent aussi leurs possibilités par des actions de groupe.
individu ne peut rien mais avec l’aide de ses congénères, il appartient alors à une force tout à fait redoutable, susceptible non seulement de s’attaquer à des ennemis en apparence bien plus importants mais également capable d’actions spectaculaires et parfois difficilement imaginables. D’autres insectes sociaux (abeilles, termites, araignées sociales, etc.) décuplent aussi leurs possibilités par des actions de groupe.
. pour en savoir plus, voir le sujet : l’intelligence animale collective
. illustration : termitière "cathédrale" au Mali
* l’espèce : le paléontologue Stephen J. Gould, souvent cité dans ces pages, va même un peu plus loin ; il pense que la sélection naturelle peut concerner des espèces entières, notamment lorsqu’elles occupent des niches écologiques voisines. Il imagine que des changements environnementaux, parfois minimes, peuvent entraîner la fusion de niches écologiques jusque là parfaitement distinctes ; de ce fait ce sont deux espèce différentes qui se disputent un même milieu et cette compétition ne finira que par la disparition de l’une des espèces concernées.
* le gène : Richard Dawkins, dans son livre « le gène égoïste » publié en 1976, prend le contre-pied des approches précédentes en allant vers l’infiniment petit, le gène, composant des chromosomes. Selon son approche, seul le gène a de l’importance et seule sa survie au fil des âges explique la sélection naturelle ; ici, le groupe n’est que le moyen de diffuser et sauvegarder les gènes et, d’ailleurs, l’individu qui les renferme dans son patrimoine génétique n’est qu’une enveloppe, un simple contenant. Bizarre à première vue, l’idée n’est plus si étrange après réflexion.
Quoi qu’il en soit et quel que soit le support ou l’aspect de l’élément à transmettre, c’est bien la sélection naturelle, indépendamment du niveau de son d’action (on peut imaginer qu’il soit multiple), qui va permettre la transmission de certains caractères aux génération suivantes. Quels en sont les principes ?
Principes de la sélection naturelle
Premier principe : la variation
Au sein d’une même population, même s’ils gardent un air « général » de parenté, les individus varient de l’un à l’autre en taille, pilosité, couleur, etc.. Sur les chromosomes des différents sujets, la diversité génétique se manifeste à l’échelle de segments d’ADN qui localement varient : mutations, recombinaison génétique (notamment par la reproduction sexuée, nous y reviendrons) voire épigénétique expliquent que les individus d’un même groupe diffèrent les uns des autres.
Deuxième principe : la sélection
Les différents individus vivent dans un environnement spécifique qui présente la particularité d’être instable et changeant sur le long terme. Parfois ces changements sont si importants (climat, phénomènes naturels, irruption brutale de prédateurs, maladies, etc.) que nombre d’individus ne possèdent pas les moyens de s’adapter. C’est à cette occasion que celui qui sera porteur d’un avantage sélectif (obtenu, par exemple, au moyen d’une mutation jusque là restée latente) prendra le dessus sur ses congénères. La conséquence en sera qu’il aura plus facilement accès à la nourriture (il restera donc en bonne santé) ou à la reproduction et aura normalement plus de descendants ce qui nécessite toutefois qu’il puisse « transmettre son avantage » d’où le troisième principe indispensable à la sélection naturelle…
Troisième principe : l’hérédité
Nous avons vu que pour qu’une population puisse s’adapter à des changements de milieu, il fallait que les individus qui la composent soient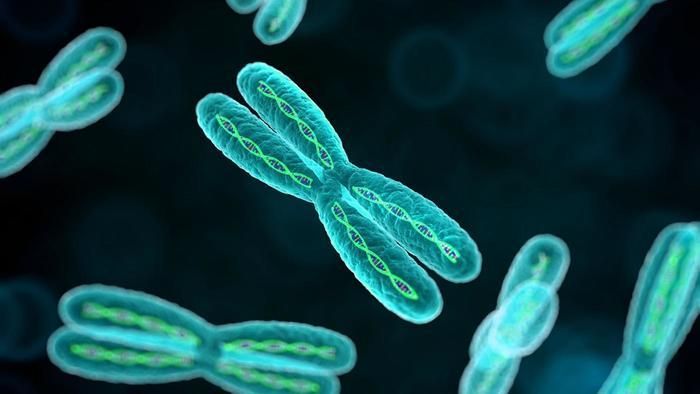 variés et que, parmi eux, les mieux armés soient sélectionnés. Encore faut-il qu’ils puissent transmettre leur héritage à leurs descendants. Si c’est le cas, l’avantage sélectif sera réparti et développé dans l’ensemble du groupe. C’est de cette façon qu’une population finit par s’adapter aux changements de milieu (en précisant toutefois qu’il ne faut pas que ce changement soit trop brutal ou trop rapide).
variés et que, parmi eux, les mieux armés soient sélectionnés. Encore faut-il qu’ils puissent transmettre leur héritage à leurs descendants. Si c’est le cas, l’avantage sélectif sera réparti et développé dans l’ensemble du groupe. C’est de cette façon qu’une population finit par s’adapter aux changements de milieu (en précisant toutefois qu’il ne faut pas que ce changement soit trop brutal ou trop rapide).
illustration : chromosomes, une approche génétique que Darwin ne pouvait connaître
De quelle façon s’exprime la sélection naturelle ?
Dans l’imaginaire du profane, compte-tenu des généralisations approximatives et des contresens parfois volontaires, la scène qui vient presque toujours à l’esprit quand on évoque la sélection naturelle est celle du lion qui course la gazelle : cette dernière sera finalement attrapée et ce d’autant qu’elle est âgée (ou trop jeune) ou malade : on imagine alors que ne subsistent chez les gazelles que les individus les plus robustes, concourant ainsi à l’amélioration de l’adaptation de l’espèce à son milieu. Stricto sensu ce n’est pas faux mais cela reste terriblement réducteur ! En effet, cet aspect « guerrier », agressif et cruel de la Nature n’existe tout simplement pas : les animaux n’ont aucune animosité les uns envers les autres, jamais de haine ou de désir de vengeance et s’ils doivent se combattre c’est parce qu’ils n’ont pas d’autre moyen pour prendre l’ascendant sur cet autre qui est en compétition avec eux. La violence pure et dure reste fréquente mais il existe bien d’autres méthodes pour circonvenir celui dont on est in fine le prédateur. Précisons d’emblée qu’il s’agit là d’agression interspécifique mais que cette compétition existe aussi au sein d’une même espèce comme nous aurons l’occasion de le voir.
* compétition directe
Il existe donc dans la Nature des couples spécifiques proie-prédateur. Ces « couples » sont effectivement très spécialisés dans la  mesure où, pour une niche écologique déterminée (et parfois même une saison précise), une proie n’a que peu de prédateurs, toujours les mêmes et qui dépendent d’elle : que la proie vienne à disparaître et le prédateur sera lui-même en grand danger, incapable le plus souvent de se fixer sur une autre proie équivalente. Prédateurs et proies ont des destins liés et la disparition de l’un peut entraîner un déséquilibre conduisant à des catastrophes écologiques. Dans la Chine de Mao, par exemple, il fut une année décidé de mettre à mort tous les oiseaux qui, semblait-il, ravageaient les vergers et diminuaient ainsi le rendement des récoltes : des dizaines de millions de volatiles furent ainsi exterminés dans l’allégresse générale. Hélas, l’année d’après il n’y eut plus de récolte du tout, les insectes, proies habituelles des prédateurs oiseaux, ayant tout dévoré.
mesure où, pour une niche écologique déterminée (et parfois même une saison précise), une proie n’a que peu de prédateurs, toujours les mêmes et qui dépendent d’elle : que la proie vienne à disparaître et le prédateur sera lui-même en grand danger, incapable le plus souvent de se fixer sur une autre proie équivalente. Prédateurs et proies ont des destins liés et la disparition de l’un peut entraîner un déséquilibre conduisant à des catastrophes écologiques. Dans la Chine de Mao, par exemple, il fut une année décidé de mettre à mort tous les oiseaux qui, semblait-il, ravageaient les vergers et diminuaient ainsi le rendement des récoltes : des dizaines de millions de volatiles furent ainsi exterminés dans l’allégresse générale. Hélas, l’année d’après il n’y eut plus de récolte du tout, les insectes, proies habituelles des prédateurs oiseaux, ayant tout dévoré.
. pour en savoir plus, voir le sujet: l’agression
. illustration : une araignée verte chasseuse d'insectes
* parasitisme
Il s’agit ici aussi d’une agression puisque la proie devient la victime de son parasite qui va profiter d’elle. Il existe néanmoins une différence capitale avec le cas précédent : autant qu’il le pourra, le parasite cherchera à être le moins agressif possible car, est-il besoin de le rappeler, la mort de la proie serait également une catastrophe pour lui, obligé de chercher à nouveau un hôte susceptible de l’héberger. C’est la raison pour laquelle certains parasites peuvent coexister étonnamment longtemps avec leurs proies, ne la quittant que lorsque celle-ci est victime d’un accident de vie, voire tout simplement de la vieillesse. Il existe évidemment diverses formes de parasitisme, depuis la guêpe fouisseuse qui pond ses œufs dans le corps d’un cafard prisonnier qui servira de réserve alimentaire pour ses larves, jusqu’à la douve du foie, parasite des ruminants dont l’homme n’est en somme qu’un hôte par défaut et même jusqu'au protozoaire qui pousse les rats au suicide…
. pour en savoir plus, voir le sujet: parasitisme et évolution
* mimétisme et discrétion
L’art de se cacher, de se fondre dans son environnement est un moyen de résister au sort contraire, un moyen de s’adapter. On cite volontiers le cas de l’ours blanc dont une mutation ancienne a permis à ceux qui en sont porteurs de mieux se fondre dans la banquise et donc d’approcher plus facilement les proies (avantage évolutif). Il existe de nombreuses formes de mimétismes, certains individus copiant à merveille ceux dont ils s’inspirent afin le plus souvent de tromper leurs propres prédateurs : ainsi, des lézards imitent à la perfection le bois de l'arbre sur lequel ils guettent (photo); ailleurs des serpents imitent la robe de plus mortels qu’eux afin de dissuader toute attaque ; ailleurs encore des orchidées imitent la forme et l’odeur de guêpes ou d’abeilles de manière à ce que l’insecte mâle trompé puisse disséminer le pollen récupéré sur son corps lors de la pseudo-copulation. Les mimétismes sont parfois si élaborés qu’on se demande quel phénomène extraordinaire a pu les produire : nul miracle n’est ici en jeu et seule, l’Évolution, par tâtonnements successifs au cours de milliers de siècles, a pu arriver à un tel résultat…
ceux qui en sont porteurs de mieux se fondre dans la banquise et donc d’approcher plus facilement les proies (avantage évolutif). Il existe de nombreuses formes de mimétismes, certains individus copiant à merveille ceux dont ils s’inspirent afin le plus souvent de tromper leurs propres prédateurs : ainsi, des lézards imitent à la perfection le bois de l'arbre sur lequel ils guettent (photo); ailleurs des serpents imitent la robe de plus mortels qu’eux afin de dissuader toute attaque ; ailleurs encore des orchidées imitent la forme et l’odeur de guêpes ou d’abeilles de manière à ce que l’insecte mâle trompé puisse disséminer le pollen récupéré sur son corps lors de la pseudo-copulation. Les mimétismes sont parfois si élaborés qu’on se demande quel phénomène extraordinaire a pu les produire : nul miracle n’est ici en jeu et seule, l’Évolution, par tâtonnements successifs au cours de milliers de siècles, a pu arriver à un tel résultat…
. pour en savoir plus, voir le sujet : le mimétisme, une stratégie d'adaptation
. pour en savoir plus, voir le sujet: comportements animaux et évolution
. illustration : geckos uroplatus, sorte de lézard mimétique
* coévolution
La coévolution est l’évolution « côte-à-côte » de deux espèces qui, dans certains cas, finissent par devenir dépendantes l’une de l’autre ; en effet, chacune des espèces trouve un bénéfice dans une collaboration qui assure à chacune survie et reproduction. Pour illustrer une coévolution d’espèces, on cite souvent l’association d’un papillon de nuit avec une orchidée de Madagascar. Cette dernière possède une extension de la corolle (pétales) appelée « éperon ». Cet organe contient le nectar mais est d’une longueur assez importante puisqu’il mesure jusqu’à 25 cm : de ce fait, sur l’île de Madagascar, seul le papillon de nuit possède une trompe assez longue pour aller puiser le nectar. Ainsi, la fleur peut se reproduire facilement grâce au papillon qui, en contrepartie, bénéficie d’une nourriture réservée. La solution trouvée ici par l’Évolution est apparemment élégante mais souffre d’un handicap certain : si l’une des deux espèces disparaît, l’autre est automatiquement condamnée.
Cas particulier de la sélection sexuelle
Cette partie du sujet a fait l’objet d’une publication spécifique (voir « reproduction sexuée et sélection naturelle »), nous n’en évoquerons donc ici que les grandes lignes.
Très tôt dans l’histoire de la théorie de l’Évolution, il avait été remarqué cette bizarrerie que semble représenter la reproduction sexuée et plus précisément la sélection sexuelle. En effet, afin de « séduire » la femelle, le mâle (c’est exceptionnellement l’inverse) est amené à prendre des risques : il peut se parer de couleurs chatoyantes  comme certains poissons ou oiseaux ou bien agrémenter sa parade du déploiement d’organes particulièrement voyants et malcommodes comme la queue du paon faisant la roue ; ailleurs, le mâle se lance dans une parade compliquée, extraordinairement codifiée, et qui demande toute son énergie car s’il manque un geste ou l’effectue improprement, la femelle se détournera de lui, persuadée qu’il ne possède pas les qualités (et les chromosomes) qu’elle recherche (exemples de nombreux oiseaux, notamment aquatiques). Dans tous les cas, le mâle fragilise sa position, soit en attirant l’attention d’un prédateur par sa nouvelle visibilité, soit en se déconcentrant et en baissant sa garde. Il doit donc bien exister un avantage quelque part pour qu'un individu se découvre ainsi puisque, au fil du temps, la sélection naturelle a conservé cette approche…
comme certains poissons ou oiseaux ou bien agrémenter sa parade du déploiement d’organes particulièrement voyants et malcommodes comme la queue du paon faisant la roue ; ailleurs, le mâle se lance dans une parade compliquée, extraordinairement codifiée, et qui demande toute son énergie car s’il manque un geste ou l’effectue improprement, la femelle se détournera de lui, persuadée qu’il ne possède pas les qualités (et les chromosomes) qu’elle recherche (exemples de nombreux oiseaux, notamment aquatiques). Dans tous les cas, le mâle fragilise sa position, soit en attirant l’attention d’un prédateur par sa nouvelle visibilité, soit en se déconcentrant et en baissant sa garde. Il doit donc bien exister un avantage quelque part pour qu'un individu se découvre ainsi puisque, au fil du temps, la sélection naturelle a conservé cette approche…
Eh bien, le risque d’être victime d’un prédateur existe mais, toutes choses pesées par ailleurs, il semble que, pour l’espèce, il soit inférieur au gain que représente le choix du meilleur mâle par la femelle. Séduire par une parade particulièrement réussie coûte peut-être la vie à quelques uns (au fond, peut-être trop hardis ?) mais permet le choix d’individus performants par rapport à ceux vécus comme plus ternes car moins entreprenants ou courageux,. Ce sont donc les chromosomes du plus vigoureux qui se distribueront plus rapidement et en plus grand nombre dans le groupe…
Nous venons d’évoquer ce que les spécialistes appellent la sélection intersexuelle, c’est-à-dire mettant en jeu mâles et femelles. Il reste à signaler un autre aspect de cette sélection par le sexe : la sélection intrasexuelle qui oppose les mâles entre eux pour la possession des femelles ; c’est, par exemple, les combats de cerfs se terminant parfois

par la mort d’un combattant (voire des deux lorsque leurs bois sont emmêlés) ou de gorilles lors de la remise en cause de la position dominante du mâle « alpha ». En terme de « rentabilité » évolutive, on peut formuler ici les mêmes remarques que pour la sélection intersexuelle.
La sélection naturelle, un item universel
La sélection naturelle est un élément majeur de la théorie de l’Évolution qui explique le fondement de l’Évolution elle-même : l’adaptabilité des espèces. Pour qu’une espèce, quelle qu’elle soit, survive au fil des centaines de milliers d’années, il lui faut s’adapter aux changements de son milieu, changements qui inéluctablement surviendront. Même ainsi, certaines transformations de la nature restent trop brutales ou trop rapides et, du coup, de nombreuses espèces disparaissent faute d’avoir pu s’adapter : ce fut le cas au cours des cinq grandes extinctions de masse qui ont peuplé l’histoire de la vie sur Terre (nous vivons d’ailleurs à cause de l’Homme actuellement une sixième extinction tout aussi terrible mais il s’agit là d’un autre sujet).
. pour en savoir plus, voir le sujet: les extinctions de masse
La vie est une course sans fin vers l’adaptation, vers une mise en conformité naturelle : le milieu change et oblige, sous peine de mort, à la mise à niveau des espèces ; la proie se dote d’une arme défensive nouvelle et oblige son prédateur à « inventer » une parade ; un nouveau prédateur apparaît qui contraint proie et prédateur de l’ancienne dualité à se « reconvertir » rapidement, etc. Il en est ainsi depuis la nuit des temps et, en dehors de toute intervention humaine, il en sera toujours ainsi. Ce paradoxe qui veut que l’on se transforme continuellement pour retrouver un équilibre antérieur se trouve résumé dans ce que l’on appelle « l’hypothèse de la reine rouge ».
Cette hypothèse de la reine rouge a été formulée par le biologiste américain Leigh Van Valen en 1972. Le biologiste fait allusion à une scène  du roman de Lewis Carol, « de l’autre côté du miroir » qui est la seconde partie, moins connue, de « Alice au pays des merveilles ». À cet instant de l’histoire, Alice se trouve sur un échiquier et est entraînée dans une course terrible par la reine rouge du jeu d’échecs ; Alice ne peut s’empêcher de demander : « mais, Reine rouge, c’est étrange, nous courons vite et pourtant le paysage autour de nous ne change pas ? » et la reine de répondre : « Nous courons pour rester à la même place ».
du roman de Lewis Carol, « de l’autre côté du miroir » qui est la seconde partie, moins connue, de « Alice au pays des merveilles ». À cet instant de l’histoire, Alice se trouve sur un échiquier et est entraînée dans une course terrible par la reine rouge du jeu d’échecs ; Alice ne peut s’empêcher de demander : « mais, Reine rouge, c’est étrange, nous courons vite et pourtant le paysage autour de nous ne change pas ? » et la reine de répondre : « Nous courons pour rester à la même place ».
Cette idée a été également reprise par l’écrivain italien Tomasi di Lampedusa dans son unique livre, « le guépard » où il fait dire à l’un de ses héros, Tancredi : « il faut tout changer pour que tout reste comme avant ! ».
La Vie est un éternel recommencement et la sélection naturelle y contribue de façon considérable.
Mise à jour 2026 : les apports récents de la biologie moderne
Depuis la rédaction initiale de cet article, plusieurs avancées importantes ont enrichi notre compréhension de la sélection naturelle. Les progrès de la génétique, de l’écologie évolutive et de la biologie des populations ont confirmé — et parfois précisé — les intuitions de Darwin.
• Le rôle central des gènes régulateurs
On sait désormais que l’évolution ne repose pas seulement sur des mutations “classiques”, mais aussi sur des modifications de l’expression des gènes. De petites variations dans les séquences régulatrices peuvent entraîner de grands changements morphologiques ou comportementaux, sans altérer les gènes eux‑mêmes.
• L’évolution rapide observée en temps réel
De nombreuses espèces montrent des adaptations mesurables sur quelques décennies seulement : moustiques du métro de Londres, lézards des îles, poissons soumis à la pêche intensive. La sélection naturelle n’est donc pas un processus lent : elle peut être fulgurante lorsque les pressions du milieu sont fortes.
• La sélection sexuelle revisitée
Les travaux récents confirment que les signaux “voyants” (plumages, parades, couleurs) sont des indicateurs fiables de bonne santé ou de robustesse génétique. La femelle ne choisit pas “au hasard” : elle sélectionne des signaux coûteux, impossibles à tricher.
• La Reine Rouge confirmée
Les études sur les parasites et leurs hôtes montrent que la coévolution permanente est un moteur puissant de diversification. Chaque espèce doit “courir pour rester à la même place”, exactement comme le suggérait Van Valen.
• Un lien direct avec les extinctions
Les crises biologiques majeures rappellent que la sélection naturelle ne suffit pas toujours : lorsque les changements sont trop rapides ou trop brutaux, même les espèces les mieux adaptées disparaissent. Ce point sera développé dans l’article consacré aux grandes extinctions (voir l'article : les extinctions de masse).


/image%2F1490480%2F20260802%2Fob_844ba8_ammonite-fossil-608164780.jpg)
/image%2F1490480%2F20260802%2Fob_99527d_zhaoianus.png)
/image%2F1490480%2F20260802%2Fob_8ca1ce_poisson-volant-exocoetidae.png)
/image%2F1490480%2F20260802%2Fob_e027db_genes-hox.jpg)



/image%2F1490480%2F20250808%2Fob_a0fc04_troupeau-elephants.jpg)
/image%2F1490480%2F20250808%2Fob_5f875b_sima-de-los-huesos-en-atapuerca.jpg)
/image%2F1490480%2F20250808%2Fob_b600ba_crane-de-benjamina-atapuerca-23622524.jpg)
/image%2F1490480%2F20250809%2Fob_3215b4_homme-neandertal-chapelle-aux-saints.jpg)
/image%2F1490480%2F20250809%2Fob_a6428c_ephedra-viridis41-4197803983.jpg)
/image%2F1490480%2F20250809%2Fob_77eb73_achillee-mille-feuilles.jpg)
/image%2F1490480%2F20250809%2Fob_9e66e0_chaman-de-bad-durrenberg.jpg)
/image%2F1490480%2F20241106%2Fob_ad4375_bocidium-globulare.jpg)
/image%2F1490480%2F20241106%2Fob_055983_mymaridae.jpg)
/image%2F1490480%2F20241108%2Fob_0b8c01_scarabee-titanus-giganteus.jpg)
/image%2F1490480%2F20241108%2Fob_d33343_mante-religieuse-1000x672c.jpg)
/image%2F1490480%2F20241108%2Fob_e439a8_pompile-vs-mygale.jfif)
/image%2F1490480%2F20241108%2Fob_881598_araignee-mymecomorphe.jpg)












































/image%2F1490480%2F20260802%2Fob_844ba8_ammonite-fossil-608164780.jpg)
/image%2F1490480%2F20260703%2Fob_2c5cf0_toiles-pop-iii-eso1524a.jpg)
/image%2F1490480%2F20260529%2Fob_972ad3_ngc-7293-helix-nebula-2.jpg)
/image%2F1490480%2F20260317%2Fob_86ec57_blouissante-galaxie-ngc-2276.jpg)
/image%2F1490480%2F20260108%2Fob_f0fb9d_gaia.jpg)
/image%2F1490480%2F20251021%2Fob_da88f0_nebuleuse-medulla.jpg)